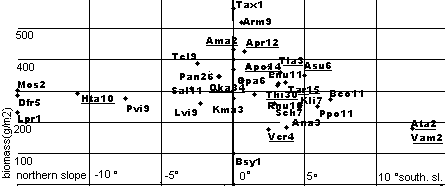
Diagram 1: Distribution of plants according to southern (positive), northern (negative) slope and above-ground biomass of surrounding grassland. Legumes are underlined.
Minor improvement potential of the vetch-rhizobia system for grassland fertility in semihumid rangelands at Xiaman, eastern edge of the Qinghai-Tibet plateau
Holger Blasum, Jiming Zhou, Ying Zeng, Shigui Liu, Denghong Zhang, Hong Xue (National Grassland Key Laboratory, Dept. of Biotechnology, Sichuan Union Univ., Chengdu 610064, PR China)
Abstract: Although the Xiaman area is diverse in wild legumes, the existence of the at least two groups of rhizobia does only lead to the formation of effective nodules in dry microhabitats (best in dunes); in plain pasture area fodder vetch was not improved by neither by inoculation with indigenous rhizobia nor with urea. Response to phosphorus seems to be more pronounced.
Introduction: Biological nitrogen fixation is the major nitrogen input pathway to natural ecosystems. There are a variety of Prokaryotes able to fix nitrogen, a relatively well-known family are the Rhizobiaceae which are symbiotic bacteria able to form nodules (small spheres) on the roots of legumes (Rao 1987). The host legumes (Fabaceae) comprise many well-known vegetables such as bean, pea or peanut as well as forages such as clover, sainfoin, vetch, milkvetch, alfalfa etc. Encouraged by a report of successful rhizobial inoculation to alfalfa in a semiarid part of the plateau (Nie 1989), the aim of this study was to assess the potential use of the rhizobia-legume biological nitrogen fixation in a state-owned sheep-raising farm in the county of Zoige/Ruoergai (33o34'N/102o28'E, 3340m a.s.l., annual mean temp. 1.1oC, annual mean precipitation 650mm, Liu 1994).
Experimental methods: (Collecting wild samples:) Soil samples and wild species of legumes had to be collected from 1m*1m squares and quantified (with special attention to nodulation patterns) by sampling along a 3-km transect crossing a northern and southern hill-slope as well as grassland of different moisture content in duplicate. Identification was done by Prof. Xu Langrang in Shaanxi and according to local herbarium specimen. All data were checked by for significance by nonparametric tests. Rhizobia were isolated from nodules according to Vincent (1970).(Investigating rhizobial physiology:) To ascertain validity of isolated strains, 54 strains of putative rhizobia of cultivated Onobrychis, Vicia sativa and wild legumes have been characterized by numerical taxonomy on 90 physiological characteristics (such as resistances, C/N source utilization, growth dynamics) and sorted by UPGMA clustering algorithm. (Field fertilization:) Using 100 miniplots in a modified Latin square design (secondary orthogonal rotated design, Chen 1986) fodder vetch seeds have been fertilized with a mixture of fermentation broths of 15 strains of indigenous rhizobia; as a control mineral fertilizers such as potassium dihydrogen phosphate (up to 75kg P/ha), urea (up to 84kg N/ha), lime (up to 750kg lime/ha), micronutrients and potassium chloride (up to 94kg K/ha)Āhave been applied in four levels.
Diagram 1: Distribution of plants according
to southern (positive), northern (negative) slope and above-ground
biomass of surrounding grassland. Legumes are
underlined.
Results: (Wild legume distribution:) A huge variety of 15 legumes has been found in a total sampling area as small as 87m2. All legumes summed up for 2.95% of total plant biomass in late August (peak point for legumes), which is in the normal range for grasslands (Zhang 1989). The distribution of the different legumes depends on soil moisture and slope angle. Legumes significantly (P<0.05) thrive best in dryer habitats, an extreme case is Astragalus sungpanensis (Asu) whichwas found in dunes. In moderately arid grasslands (with high evaporation such as southern slopes or sandy soils, the sedge Kobresia maquensis (Kma) occurs) other Astragali as well as the grazing-resistant Trigonella archiducis-nicolai (Tar) and stout Thibetia himalaica (Thi) dominate, whereas Oxytropis kansuensis (Oka)(the most common legume) and Hedysarum (Hta) extend to slightly moister areas; in really damp swamplands no legumes exist. Nodule color usually was rather rose to red in dry and sandy environments and white or brown in moister environments.
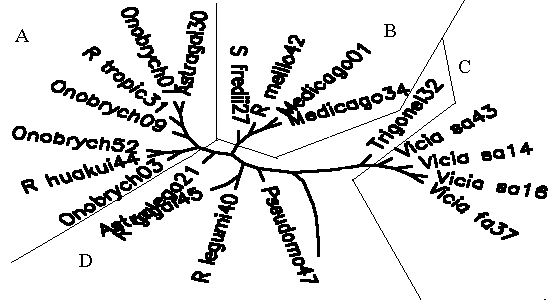
Diagram 2:Āupper part of dendrogram
(bearing putative rhizobia) generated by modified chisquare
coefficient, A is R. huakuii cluster, B is
Sinorhizobium cluster, C is vetch, D is mixed and outgroup
branch.
(Rhizobial classification:) By using a chi-square-based semimodified similarity coefficient, after removing strains growing at 4oC (freezer-selected contaminants), bacteria isolated from Ruoergai/Hongyuan vetch clustered with isolates from subtropical (Chengdu) vetches (C area), on the other hand an assemblage of R. huakuii type strain, medic, Onobrychis and Astragalus spp. strains forms a second cluster (A-B area). C differs from (A-B) in that the vetch cluster is resistant to 2000ppm manganese, cannot use citrate, is susceptible to 5000ppm potassium sulfate and susceptible to 4-10ppm malachite green. Some medic-derived isolates form a strong subcluster (B)Āwithin the second (A-B)Ācluster. (Field fertilization:) Inoculating vetch with rhizobia led to a very remarkable (P<0.0001) change in nodulation pattern: inoculated vetches are much earlier nodule-bearing than control plants (the crown root is more nodule-bearing than with controls). However this increased crown root nodulation has led to a decrease in nodulation in the side-roots, so that overall nodulation changed very little. Urea fertilization, micronutrients and liming failed to raise any growth parameters, whereas potassium dihydrogen phosphate dressing made the DM above-ground total yield go up by 60.0% (P<0.05), nodule weight increased by 146% (P<0.002). Potassium chloride alone seemed also powerful (+17.1% and +62.5%), but due to a low number of plots (4 ) results are not statistically significant. The striking effect of phosphorus fertilization corresponds to the finding that the relation of grassland productivity to rapidly available phosphorus was more inversely related (P<0.0012) than the inverse relation of grassland productivity to alkali-digestible nitrogen (P<0.13).
Discussion: On the experimental site fodder vetch yield was neither increased by rhizobial inoculation nor urea. The altered nodulation pattern demonstrated that at least for vetch group rhizobia a plant nodulation autocontrol mechanism (Caetano-Gresshoff 1991) was invoked. One may argue this may be different with sainfoin or medic which apparently belong to another rhizobium (cross-inoculation?) group. This view could be backed by the observation that natural nodulation of sainfoin and medic is poorer than that of vetches. However even for oat and weeds urea response is relatively little (6kg and 2.3kg above-ground DM increase per kg urea), so that Wang's (1994) statement that in N-rich parts of the Qinghai-Tibet plateau N-fertilization is relatively inefficient applies. Again it is emphasized that rhizobial nitrogen fixation as well as the usefulness of host plants vary between microclimates: In semimoist grassland where Oxytropis dominates, white nodules indicate low fixation efficiency , and the absence of legumes in equally productive moister communities indicates that BNF is probably amply provided by other symbioses, such as arctic-sedge-associated free-living baxcteria (Nosko 1994) and Cyanobacteria (Henry 1986, Hendriksson 1987), furthermore Oxytropis kansuensis contains the poisonous swainsonine (Lu 1993). On the other hand, in the disturbed and dry plots nodules were pink and thus leghemoglobin-containing indicating active N-fixation, in the dunes legumes even accounted for 11.6% of biomass and here the legume-rhizobia system might prove very valuable for land reclamation, as legumes are known to be good primary successors (Kenny 1990, Blundon 1990).
References:Ā